Biological Control of Weeds: Effects of Rhizobacterial Inoculation
on the Growth of Velvetleaf (Abutilon theophrasti
Medik.)
Maria Fatima T. Begonial , Robert J. Kremer2,
and Gregorio B. Begonial
lDepartment of Biology, Jackson State University, Jackson, MS 39217,
and 2USDA-ARS, University of Missouri, Columbia, MO 65211
Preemergence application to soil of rhizobacteria as biological agents in
controlling weeds in the growth chamber, greenhouse and field were carried
out to evaluate their effectiveness. Results indicated that various
rhizobacterial isolates inhibited weed seedling growth and exhibited deleterious
effects including stunted, necrotic tap roots, severely inhibited lateral
root development and chlorotic foliage. Soil inoculation studies with selected
rhizobacteria resulted in reduced seedling emergence, decreased seedling
vigor, and severely damaged root systems. Results also indicated that
rhizobacteria can be effective biocontrol agents at the rate of
108109 cells in the growth chamber, greenhouse
and field whether they are used either as liquid or peat-based inoculants.
Velvetleaf is a very competitive, annual, dicotyledonous plant that is a major weed problem in row crops grown in the United States (Spencer, 1984). It produces high numbers of seeds, many of which are dormant, and after dispersal remain in soil for several years during which germination gradually occurs (Egley and Chandler, 1983).
Several preemergence and postemergence herbicides can control velvetleaf but control levels have been inconsistent over several years. General recommendations for chemical control of velvetleaf can not be made because of differences in climate, soils, crop rotations and cultural practices (Spencer, 1984). Some widely used herbicides have been implicated in contamination of ground water, soils, and food products, which may threaten public health and safety. Because of the public concern about environmental safety of herbicides, interest in developing effective nonchemical weed management methods has increased (Kremer and Kennedy, 1996).
A potential method of controlling weeds is the use of microorganisms (Rice, 1974). Rhizobacteria are attractive for biocontrol because the seed and/or seedling is attacked rather than the growing plant. To be effective biological agents, the microorganisms should have the following major characteristics (Begonia et al., 1990; Begonia and Kremer, 1994). The rhizobacteria should be: (a) selectively attracted to weed seedlings, (b) toxic or be able to infect their host plants resulting in seed or seedling diseases, and (c) able to rapidly colonize seeds or roots of weed species resulting in seed decay, inhibition of seedling emergence, and decreased seedling vigor. The examination of microorganisms associated with weed seeds and roots (Kirkpatrick and Bazzas, 1979; Mishra and Srivastava, 1977, Kremer et al., 1990) has been reported. The use of fungi (mycoherbicides) as biological agents in controlling weeds is well documented (Templeton et al., 1979). The existence of deleterious rhizobacteria on the roots of weed seedlings has been reported for velvetleaf (Kremer et al., 1990) and for downy brome (Cherrington and Elliott, 1987).
Although rhizobacteria show potential as herbicide, little information is available regarding the root colonization capacity as well as the effects of these rhizobacteria on plant growth. Research is needed regarding formulation rates and application timing to control weeds most effectively and that could be integrated into existing agricultural practices.
The objective of this investigation was to evaluate the effectiveness of
preemergence applications of rhizobacteria as biological agents in controlling
weeds when applied as liquid inocula to soil under growth chamber, greenhouse
and field conditions. Birdsfoot trefoil was included in this study as a
representative domestic plant having similar characteristics of many weed
species.
MATERIALS AND METHODS
Growth chamber experiments
Plant host--A plant accession of velvetleaf seeds (Mississippi, 1985) was used as the host plant of the rhizobacteria isolated from velvetleaf. Seeds were surface-sterilized by immersion in 1.25% (w/v) sodium hypochlorite for 8 min, followed by immersion in 70% ethanol (v/v) for 4 min. The seeds were washed thoroughly (at least ten times) with sterile distilled water and allowed to imbibe water for 24 hrs at 27C. Seeds with emerging radicles (~2 mm) were used in inoculation experiments which examined the development of pure cultures at several concentrations on growth of the microbes in soil during a two-week growth period of the test plants.
Soil materials--The field soil (Mexico silt loam) used in the experiments had a pH of 5.9; 1.95% organic matter; 20 kg/ha available P; 3,370 kg/ha Ca; 438 kg/ha Mg and 128 kg/ha K. Soils were sieved through a number 6 grid aluminum screen to remove clods, roots and stones. Three hundred fifty gram lots of soil were placed in autoclavable polyethylene plastic bags and sterilized by autoclaving at 103.5 kPa for 1 hr on three successive days. Freeze-dried cells resuspended in phosphate buffered saline (PBS) were incorporated and mixed thoroughly with the soil to yield final densities of 107, 108, and 109 cells/g dry soil. The inoculated soil in each bag was transferred into disinfected pots.
Bacterial cultures and culture conditions--Rhizobacterial isolates from velvetleaf, VLBR 01 and V 239, were cultured in King's broth (Sands and Rovira, 1970) and shaken at 27C for 27 hrs. The broth cultures had a pH of 7.2. Cells were harvested by centrifugation (6,000 rpm) at 5C and then washed three times with PBS (pH 7.4). Concentrated cells were suspended in a sterile-freeze drying suspension medium composed of 7.5 g sucrose, 5.0 g dextrin, and 1.0 g sodium glutamate in 100 ml distilled water and freeze-dried. Freeze-dried cells were resuspended in PBS to yield the desired cell concentrations. The number of cells present in the inoculant was determined by spread plating appropriate dilutions of serially diluted bacterial cell suspensions on King's agar.
Inoculation experiments--Ten surface-sterilized, pregerminated (~2 mm radicles) seeds were planted in each pot following the procedures of Scher et al. (1984). The plastic pot used was 9.0 cm. high, with a top and bottom diameter of 12.0 cm and 7.0 cm, respectively. Each pot received 20 ml of plant nutrient solution supplemented with 1.5 mM KNO3 (Hoagland and Arnon, 1938). Each of the bacterial cultures (VLBR 01 and V 239) was inoculated into the soil at three cell concentrations (107, 108 and 109 cells/g dry soil) and each was replicated three times. Noninoculated soil planted with seeds served as controls (CON). The seedlings were grown at 28C during a 16 hr light period and at 21C during an 8 hr dark period. Light was supplied by fluorescent and incandescent lamps at a photon flux density of 230250 µmol m-2 s-1. The seedlings were grown for 2 weeks in the growth chamber. The growth chamber experiments were set up in completely randomized design with three replications per treatment.
Data collection and analysis--At harvest, rhizosphere
soil (that which remained clinging to the roots when plants were pulled from
the pots) was separated from root systems by immersion in PBS diluent of
the corresponding rhizosphere soil (same treatment). Roots that had been
freed of soil and non-rhizosphere soil were also suspended separately in
PBS containing 0.01% v/v Tween 40 and agitated vigorously on a vortex shaker
for 5 min. Bacterial populations were determined by serially diluting (10-fold
steps) the root, the rhizosphere soil and the non-rhizosphere soil suspensions
in PBS blanks and spreading onto duplicate plates of King's agar medium.
Bacterial colonies were enumerated after incubation at 27C for 72 hrs. The
length of the primary tap root was measured before discarding. The wet and
dry weights of the rhizosphere and non-rhizosphere soils were taken before
and after drying them in the oven at 105C for at least 24 hrs. To determine
the effect of rhizobacterial inoculation on the growth of velvetleaf, plant
tops or shoots were also harvested, dried at 65C for at least 24 hrs and
shoot dry weight of plants per treatment was determined. Visual observations
of gross root morphology were also made. All data were subjected to analysis
of variance and treatment means were compared using the LSD test at the 5%
level of probability.
Greenhouse experiments
Plant hosts--Seeds of velvetleaf (Mississippi, 1985) and birdsfoot trefoil (MU 81), were surface-sterilized as before, and the growth of two-week old inoculated and noninoculated seedlings were evaluated as previously described in the growth chamber experiments.
Soil materials--Soil was prepared as described
for the growth chamber experiments except that liquid inoculants were
incorporated and mixed thoroughly with the soil to yield final densities
of 109 cells/g dry soil. The inoculated soil in each bag was
transferred into disinfected pots. Inoculants consisted of antibiotic resistant
strains to allow for their identification in the soil after the prescribed
period of growth.
Screening rhizobacterial isolates for resistance to antibiotics
Antibiotics--Nalidixic acid (NA), rifamficin (RIF), streptomycin sulfate (STR) and tetracycline (TET) were obtained from Sigma Chemical Company, St. Louis, MO. Stock antibiotic solutions were prepared according to Kremer and Peterson (1982). Specifically, STR and RIF were prepared in 100 ml distilled water while TET and NA were dissolved in small volumes of 95% ethanol and 0.1% NaOH, respectively, before addition of distilled water. The stock solutions were filter-sterilized through Gelman filter assemblies (0.2 µm) attached to a syringe into sterile bottles and stored at 4C. Fresh stock solutions were prepared before each test. Antibiotic supplemented King's agar media were prepared by dispensing the appropriate volume of stock antibiotic solution into sterile petri dishes, followed by the addition of autoclaved King's agar cooled to 50C.
Screening procedure--Wild-type rhizobacteria (VLBR 01, V 239, LCBR 04, LCBR 07 and 22411 SSDW) were streaked onto antibiotic test plates to determine the intrinsic antibiotic resistance levels for each isolate. The test plates contained a range of concentrations of each antibiotic on individual King's agar plates. Antibiotic plates were incubated at 27C for 3 days and 7 days or longer for fast and slow growers, respectively. The growth on the antibiotic plates was compared with that in King's agar control plates and scored as positive or negative.
Bacterial cultures and culture conditions--Rhizobacterial isolates VLBR 01 and VLBR 07 from velvetleaf; LCBR 04 and LCBR 07 from birdsfoot trefoil and a soil bacterium 22411 SSDW were used for inoculation experiments. Antibiotic resistant bacterial isolates were cultured in King's broth (Sands and Rovira, 1970) shaken at 27C for 72 hrs. The broth cultures had a pH of 7.2. Cells were harvested by centrifugation (6,000 rpm) at 5C and then washed three times with PBS (pH 7.4). Concentrated cells were resuspended in PBS yielding the desired cell concentrations. The number of cells present in the inoculant was determined by spread plating appropriate dilutions of serially diluted bacterial cell suspensions on King's agar.
Inoculation experiment--Procedures for this experiment are similar to those protocols described in the growth chamber experiments.
Data collection and analysis--Data collection and
analysis were done as previously described in the growth chamber experiments.
Field microplot studies on selected rhizobacteria
Screening rhizobacterial isolates for resistance to antibiotics--Screening of rhizobacterial isolates for resistance to antibiotics was carried out as described in the greenhouse experiments.
Inoculum production--Rhizobacterial isolates VLBR 01 and V 239 and LCBR 04 and LCBR 07 isolated from velvetleaf and birdsfoot trefoil seedling roots, respectively were used as inoculants. Cells of each bacterial isolate were produced by inoculating a loopful of each test culture into 250 ml Erlenmeyer flasks each containing 125 ml of King's broth which were then incubated on a rotary shaker for 4872 hrs at 150 rpm at 28C. The cells were harvested by centrifugation at 6,000 rpm for 20 min at 5C and washed three times with phosphate buffered saline (PBS, 0.01 M K2HPO4, 0.14 M NaCl, pH 7.2). Pelleted cells were resuspended in minimal volume of PBS and maintained at 8C. Counts of concentrated cells for each culture were determined by spreading 0.1 ml samples of appropriate dilutions on duplicate King's agar plates. Plates were incubated at 27C for 23 days after which colonies were counted.
Preparation of inoculum--Two hundred gram quantities of granulated peat were dispensed into each autoclaved plastic bag after neutralizing the peat to pH 6.87.0 with CaCO3. The neutralized peat was sterilized by autoclaving for 15 min at 120C at 103.5 kPa pressure for three successive days. The sterilized peat was inoculated with 80 ml of each cell suspension containing 109 cells/ml based on plate count. Bags of inoculated peat were incubated aseptically for 35 days at 25C. The number of cells/g of peat was determined by plate count as previously mentioned. Packaged inocula were stored at 8C until ready for use.
Field site and experimental design--Applications of peat inoculants containing rhizobacteria were made on each test plot (Mexico silt loam soil) measuring 1 x 1 m separated by 1 m alleys prior to velvetleaf emergence. The test was arranged as a randomized complete block with three replications.
Inoculum application--Each peat inoculant treatment (25 g) was broadcast by hand to microplots before planting. This was equivalent to 250 kg/ha rate, which approximated a rate of 2.5 x 1013 cells per ha. The inoculant was raked into the upper 45 cm of the soil immediately after application. Application was done during the cool part of the day.
Seeding--Each plot was planted with 100 velvetleaf or birdsfoot trefoil seeds sown 23 cm deep in 2 rows 30 cm apart (50 seeds/row). Control plots received noninoculated sterilized peat while background plots received no seeds and no inoculant. Plots were watered after sowing to ensure seedling emergence and were watered thereafter as needed.
Data collection and analysis--All data were collected
2 weeks after planting. Seedling emergence, seedling vigor (approximate height,
chlorosis and necrosis) and root ratings were determined. Rhizosphere
colonization by each of the bacterial isolates was determined by plating
the roots plus adhering soils on King's agar amended with antibiotic
concentrations appropriate for each isolate. All data were subjected to analysis
of variance and treatment means were compared using the LSD test at the 5
% level of probability.
RESULTS AND DISCUSSION
Various effects of rhizobacteria on velvetleaf seed germination and seedling
growth in sterile soil were observed under growth chamber conditions
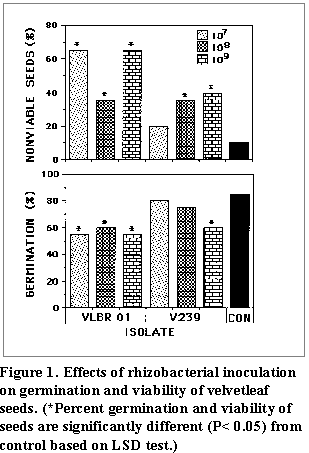
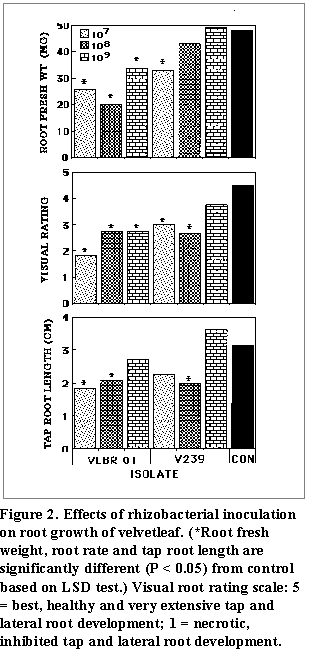
(Fig. 1 and Fig. 2). At all
rates, rhizobacterial isolate VLBR 01 significantly increased nonviable seeds
when compared to the control (Fig. 1). Correspondingly,
VLBR 01 significantly reduced seed germination compared to the noninoculated
treatment (Fig. 1). Plants inoculated with VLBR 01
had shortened tap root systems (Fig. 2). In control
plants, lateral roots were abundant and the foliage developed a dark green
color. Isolate V 239 exerted somewhat deleterious effects at the same
concentrations but results obtained were more inconsistent than those obtained
with VLBR 01. The deleterious effects of rhizobacteria on seeds, shoot and
root morphogenesis may be related to the production of phytotoxins by
rhizobacteria. Isolates used in this study were gram negative, oxidase negative,
fluorescent rods closely related to many Pseudomonas phytopathogens
which may produce toxins during root colonization thereby reducing plant
growth (Suslow and Schroth, 1982). Preliminary laboratory experiments (Kremer
et al., 1989) indicated that these rhizobacterial isolates are phytopathogenic
based on Escherichia coli inhibition tests (Gasson, 1980) and
phytotoxicity assays using cell-free filtrates of selected deleterious
rhizobacteria (Kremer et al., 1989). Results from growth chamber experiments
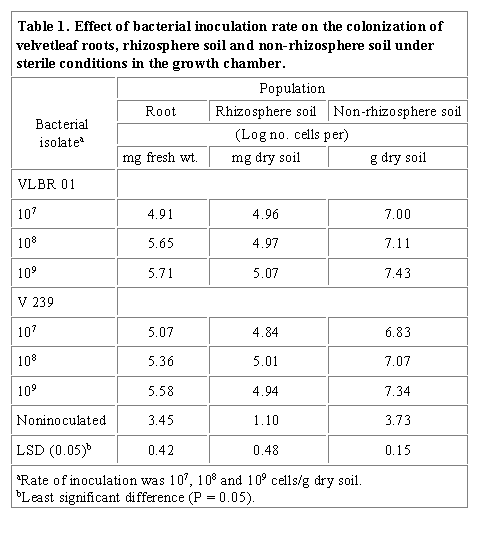
also indicated that rhizobacterial isolate VLBR 01 and V 239 attained
significantly higher populations on roots, in the rhizosphere and non-rhizosphere
soil than the same sites for noninoculated seedlings
(Table 1). Since sterile soil was used in this
experiment, it is assumed that bacteria detected in the noninoculated seedlings
originated from the seeds. Thus, it appears that all three sites may be colonized
by rhizobacteria. The root and rhizosphere soil samples were colonized by
a higher number of bacteria than the root-free soil. These results agree
with the work of Rouatt and Katznelson (1961) and Rovira and Davey (1974)
wherein they reported that the rhizosphere of crop plants contains higher
bacterial counts than the root-free soil.
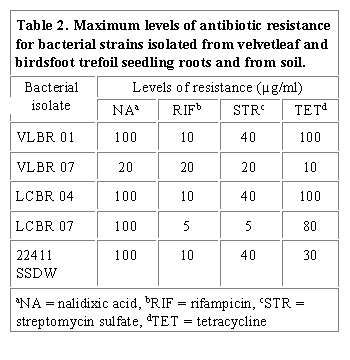
Table 2 shows the maximum levels of antibiotic resistance determined for bacterial isolates from velvetleaf and birdsfoot trefoil seedling roots and from soil. The highest levels of resistance occurred with nalidixic acid for all the rhizobacteria, except VLBR 07. VLBR 01 and LCBR 04 also exhibited higher levels of resistance to tetracycline than LCBR 07, VLBR 07 and 22411 SSDW. For example, VLBR 01 and LCBR 04 were resistant to nalidixic acid and tetracycline at 100 µg/ml and LCBR 07 was resistant to the same antibiotics at 100 and 80 µg/ml, respectively. Although 22411 SSDW exhibited high level of resistance to nalidixic acid (100 µg/ml), its resistance to tetracycline was low (30 µg/ml). Resistance to the other antibiotics varied among bacterial isolates. The above antibiotic resistance patterns specifically to tetracycline, which we observed, contradict reports for rhizobia (Schwinghamer, 1967; Davis, 1962; Pugashetti and Wagner, 1980), which were very sensitive to tetracycline. Our results agree somewhat with the results of Pankhurst (1977) who reported that some strains of Lotus rhizobia possessed high levels of resistance to tetracycline. In general, however, our results support the suggestion of Kremer and Peterson (1982) and Hagedorn (1979) that generalizations regarding the antibiotic resistance patterns of different strains or types of bacteria, specifically Rhizobium, are often invalid. The intrinsic antibiotic resistance markers selected for differentiation of inoculant strains in roots, rhizospheres and in root-free soil of greenhouse plants are shown in Table 3. Combinations of resistance to the four antibiotics, nalidixic acid, rifamficin, streptomycin and tetracycline were used to identify the inoculant strains.
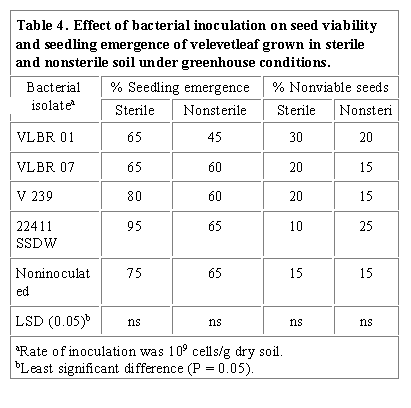
Results obtained from greenhouse evaluations of the effects of selected rhizobacteria and a soil bacterium on velvetleaf and birdsfoot trefoil seedling growth are shown on Tables 4, 5 and 6. Rhizobacterial isolates VLBR 01 increased the nonviability of velvetleaf seeds while other isolates exerted variable effects in both soil treatments compared to the controls (Table 4). VLBR 01 and other isolates decreased seedling emergence although not significantly different from the control. V 239 and 22411 SSDW slightly increased percent seedling emergence in sterile soil while no significant effect was observed in nonsterile soil. Noninoculated seedlings grown in sterile and nonsterile soil exhibited vigorous growth indicated by dark green foliage. In contrast, the inoculated seedlings exhibited chlorotic foliage first developing 710 days past inoculation. Seedlings inoculated with VLBR 01 had significantly shorter tap roots and lower root fresh weight compared to controls whether plants were grown in sterile or nonsterile soil (Table 5). Seedlings inoculated with VLBR 07 and 22411 SSDW had significantly shortened tap roots when grown in nonsterile soil but in sterile soil root length was not different from controls. All isolates significantly decreased the root fresh weight of inoculated seedlings in both soil treatments (Table 5). Rhizobacterial isolate V 239 significantly increased the shoot dry weight of inoculated seedlings in both soil treatments while 22411 SSDW exerted similar effect only in sterile soil. VLBR 01 and VLBR 07 had no significant effect on shoot dry weight of inoculated seedlings in both soil treatments (Table 5).
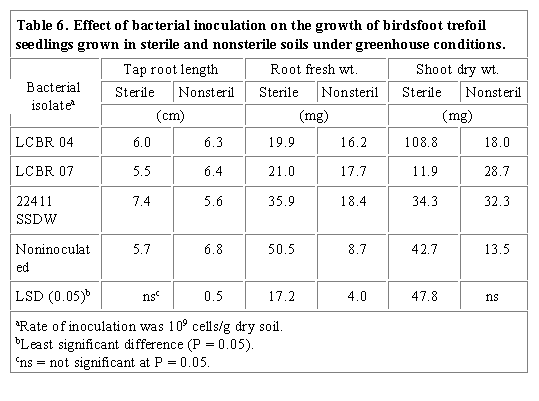
Birdsfoot trefoil seedlings inoculated with rhizobacterial isolates LCBR 04 and LCBR 07 had shorter tap roots which were not significantly different from controls in both soil treatments (Table 6). Both isolates significantly decreased root fresh weight of inoculated seedlings grown in sterile soil compared to controls. Both isolates and 22411 SSDW significantly increased the root fresh weight of seedlings grown in nonsterile soil compared to controls. Shoot dry weights were variable indicating that this parameter may not be a good indicator of deleteriousness of rhizobacterial isolates compared to root measurements.
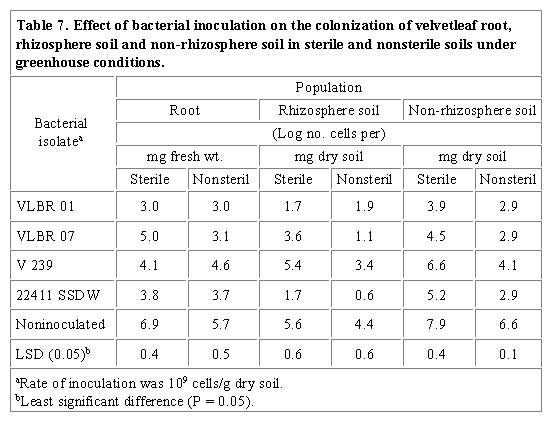
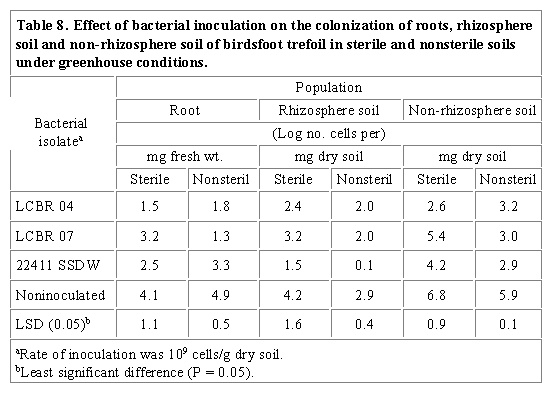
Similar results had been shown for wheat and barley where shoot chlorosis was caused by rhizosphere pseudomonads associated with their roots (Bennett and Lynch, 1981). Foliage chlorosis and root morphogenesis may be related to phytotoxins produced by rhizobacteria (Suslow and Schroth, 1982). Inhibition of root growth and development in sprouting canola (Brassica campestris L. 'Candle') (Campbell et al., 1987) and in the weed downy brome (Bromus tectorum L.) (Cherrington and Elliott, 1987) by several pseudomonads have been observed. The increased root fresh weight of birdsfoot trefoil seedlings grown in nonsterile soil (Table 6) may be attributed to plant growth promoting effect of other more competitive organisms than the inoculant strains. Inhibition tests of bacteria cultured from the root, rhizosphere and non-rhizosphere soil of birdsfoot trefoil seedlings on inoculant rhizobacteria indicated that a total of 86%, 72% and 57 % of the subcultures inhibited LCBR 04, LCBR 07 and 22411 SSDW, respectively. The increased shoot dry weight of inoculated velvetleaf and birdsfoot seedlings grown in sterile soil may also be explained by plant growth promoting microorganisms possibly originating from velvetleaf and birdsfoot trefoil seeds. Plate counts on antibiotic amended media of roots, rhizosphere and root-free soils of velvetleaf and birdsfoot trefoil seedlings grown under greenhouse conditions showed that recovery of all inoculant strains from both plant species was higher from those grown in sterile than nonsterile soil (Tables 7 and 8). Low recovery of inoculant strains from nonsterile soil may be due to competitive effects of other microorganisms colonizing the plant root surface, rhizosphere and non-rhizosphere soil under field conditions. Indeed, antibiosis assays revealed that 122%, 10% and 43% of the total bacteria subcultured from the root, rhizosphere and non-rhizosphere soil of velvetleaf, respectively, inhibited VLBR 01, VLBR 07, V 239 and 22411 SSDW. For birdsfoot trefoil, 85.5%, 71.5% and 57.2% of the bacterial subcultures inhibited LCBR 04, LCBR 07 and 22411 SSDW, respectively. Data also indicate that the velvetleaf root surface was colonized by a higher number of bacteria than the rhizosphere and non-rhizosphere soil (Table 7) in both soil treatments, suggesting that the roots may be selectively colonized by the bacterial isolates. This trend was not observed, however, in birdsfoot trefoil as the non-rhizosphere soil was colonized by a higher number of bacteria than the root surface and rhizosphere soil (Table 8). It is possible that soilborne, seedborne or airborne bacteria were present that exhibited the same multiple intrinsic antibiotic resistance patterns as that of the birdsfoot trefoil inoculants. Another explanation for the higher colonization of the root-free soil is that inoculants mixed thoroughly with soil used in each pot before sowing multiplied more rapidly in this site compared to the root surface and rhizosphere of the birdsfoot trefoil. Emergence of birdsfoot trefoil was considerably slower than velvetleaf, which may have allowed inoculant rhizobacteria to establish in soil.
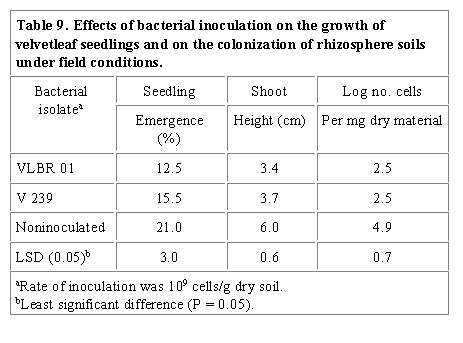
Data obtained from preliminary field evaluation of preemergence applications
of rhizobacterial isolates VLBR 01 and V 239 applied as granular (peat-based)
inocula to soil indicated that both inoculants significantly decreased seedling
emergence and shoot height of inoculated velvetleaf plants compared to
noninoculated controls (Table 9).
Table 9 also shows that a high number of the native
bacterial strains possessed the same antibiotic resistance patterns as the
inoculant strains as indicated by a high number of cells obtained from
rhizospheres of noninoculated controls (4.99) compared to those inoculated
with VLBR 01 and V 239 (2.46 and 2.47). This may have been due to a highly
competitive native bacterial population, which allowed more chance of similar
patterns occurring in the native strains. Although a high number of bacteria
with similar antibiotic patterns colonized the rhizosphere of noninoculated
controls, these bacteria may not have been as effective as the inoculant
strains. VLBR 01 and V 239, although low in population were significantly
detrimental to the velvetleaf seedlings. Both isolates may have been very
competitive during which phytotoxins detrimental to the seedlings were produced
(Suslow and Scroth, 1982). Since both inoculant strains produced fluorescent
pigments on King's agar, the possibility exists that both isolates produced
fluorescent siderophores that complexed iron in the rhizosphere thus depriving
pathogens or other members of the soil microflora of available iron necessary
for growth. This could result in a better environment for both rhizosphere
multiplication, colonization and phytotoxin production by both rhizobacterial
inoculants. The production of fluorescent siderophores has been reported
only for plant growth promoting rhizobacteria (Kloepper, 1983; Kloepper et
al., 1983). However, production of such compounds by rhizobacteria in
situ has not been reported. The peat carrier may have protected the
rhizobacteria from adverse environmental conditions enhancing the herbicidal
activity. Alginate granules have been reported as effective formulations
for mycoherbicides (Walker and Connick, 1983). However, the use of peat in
the formulation of bacterial herbicides targeted at weed seeds and seedlings
has not been reported. No field data was obtained for birdsfoot trefoil due
to poor emergence, which may be attributed to very hot and dry weather
conditions.
SUMMARY AND CONCLUSIONS
This study demonstrates that certain rhizobacteria from velvetleaf are
potentially useful in suppressing weed growth. These rhizobacteria applied
as liquid or peat-based inoculants can be effective biocontrol agents under
growth chamber, greenhouse and field conditions. Commercial use of rhizobacteria
as a herbicide requires further selection of strains to ensure consistent
weed control. Improvements in biocontrol of weeds must be achieved by selecting
rhizobacterial strains that are particularly deleterious or antagonistic
to the target weed but not to the crop. It would also be necessary to maintain
high populations of introduced rhizobacteria in proximity to the seed, seedling
root or soil. At least 108109 colony forming
units per seed or per gram of soil was used in this study for significant
reduction of weed seed viability and seedling vigor. Although these preliminary
experiments gave promising results, challenges remain in developing an effective
biological control agent for weeds. One of the most important challenges
is the elucidation of environmental factors influencing growth, survival
and colonization of rhizobacteria and also the basic mechanisms underlying
their antagonistic interaction with the weed host plants. Therefore, further
field studies are required to determine the adaptability and persistence
of applied rhizobacteria under a wide range of soils, crop and weed varieties
and environmental conditions.
ACKNOWLEDGMENTS
We thank L. Stanley for her technical assistance and the Cropping Systems
and Water Quality Research, ARS-USDA for financial support.
LITERATURE CITED
Begonia, M.F.T., and R.J. Kremer. 1994. Chemotaxis of deleterious rhizobacteria to velvetleaf (Abutilon theophrasti Medik.) seeds and seedlings. FEMS Microbiol. Ecol. 15:227236.
Begonia, M.F.T., R.J. Kremer, L. Stanley, and A. Jamshedi. 1990. Association of bacteria with velvetleaf roots. Transactions Missouri Acad. Sci. 24:1726.
Bennett, R.A., and J.M. Lynch. 1981. Bacterial growth and development in the rhizosphere of gnotobiotic cereal plants. J. Gen. Microbiol. 125:95102.
Campbell, J.N., D.D. Cass, and D.J. Peteya. 1987. Colonization and penetration of intact canola seedling roots by an oppportunistic fluorescent Pseudomonas sp. and the response of host tissue. Phytopathology 77:11661173.
Cherrington, C.A., and L.F. Elliott. 1987. Incidence of inhibitory pseudomonads in the Pacific Northwest. Plant Soil 101:159165.
Davis, R.J. 1962. Resistance of rhizobia to antimicrobial agents. J. Bacteriol. 84:187188.
Egley, G.H., and J.M. Chandler. 1983. Longevity of weed seeds after 5.5 years in the Stoneville 50-year buried seed study. Weed Sci. 31:264270.
Gasson, M.J. 1980. Indicator technique for antimetabolite toxin production by pathogenic species of Pseudomonas. Appl. Environ. Microbiol. 39:2529.
Hagedom, C. 1979. Relationship of antibiotic resistance to effectiveness in Rhizobium trifolii populations. Soil Sci. Soc. Am. J. 43:921925.
Hoagland, D.R., and D.I. Arnon. 1938. The water culture method for growing plants without soil. Calif. Agric. Exp. Sta. Cir. 347.
Kirkpatrick, B.L., and F.A. Bazzas. 1979. Influence of certain fungi on seed germination and seedling survival of four colonizing annuals. J. Appl. Ecol. 16:515527.
Kloepper, J.W. 1983. Effect of seed piece inoculation with plant growth promoting rhizobacteria on population of Erwinia carotovora on potato root and daughter tubers. Phytopathology 73:217219.
Kloepper, J.W., J. Leong, M. Teintze, and M.N. Scroth. 1980. Enhanced plant growth by siderophores produced by plant growth promoting rhizobacteria. Nature 286:885886.
Kremer, R.J., and H.L. Peterson. 1982. Nodulation efficiency of legume inoculation as determined by intrinsic antibiotic resistance. Appl. Environ. Microbiol. 34:636642.
Kremer, R.J., and A.C. Kennedy. 1996. Rhizobacteria as biological control agents of weeds. Weed Tech. 10:601609.
Kremer, R.J., M.F.T. Begonia, L. Stanley, and E.L. Lanham. 1990. Characterization of rhizobacteria associated with weed seedlings. Appl. Environ. Microbiol. 56:16491655.
Kremer, R.J., M.F.T. Begonia, and L. Stanley. 1989. Deleterious rhizobacteria for biocontrol of weed seeds and seedlings in soil. The Rhizosphere and Plant Growth Beltsville Symposium XIV Abst., p. 67.
Mishra, R.R., and V.B. Srivastava. 1977. Comparison of microflora associated with certain crops and weed seeds. Acta Mycol. 13:145149.
Pankhurst, C.E. 1977. Symbiotic effectiveness of antibiotic-resistant mutants of fast- and slow-growing strains of Rhizobium nodulating Lotus species. Can. J. Microbiol. 23:10261033.
Pugashetti, B.K., and G.H. Wagner. 1980. Survival and multiplication of Rhizobium japonicum strains in silt loam. Plant Soil 56:217227.
Rice, E. L. 1974. Allelopathy. Academic Press, Inc., New York.
Rouatt, J.W., and H. Katznelson. 1961. A study of the bacteria on the root surface and in the rhizosphere soil of crop plants. J. Appl. Bacteriol. 24:164171.
Rovira, A.D., and C.B. Davey. 1974. Biology of the rhizosphere. Pages 153204 in E.W. Carson, ed., The plant root and its environment. University of Virginia Press, Charlottesville.
Sands, D.C., and A.D. Rovira. 1970. Isolation of fluorescent pseudomonads with selective medium. Appl. Microbiol. 20:513514.
Scher, F.M., J.S. Ziegle, and J.W Kloepper. 1984. A method for assessing the root colonizing capacity of bacteria on maize. Can J. Microbiol. 30:151157.
Schwinghamer, E.A. 1967. Effectiveness of Rhizobium as modified by mutation for resistance to antibiotics. Antonie van Leeuwenhoek J. Microbiol. Serol. 33:121136.
Spencer, N.R. 1984. Velvetleaf Abutilon theophrasti (Malvaceae), history and economic impact in the United States. Econ. Bot. 38:407416.
Suslow, T.V., and M.N. Schroth. 1982. Role of deleterious rhizobacteria as minor pathogens in reducing crop growth. Phytopathology 72:111115.
Templeton, G.E., D.O. Teebest, and R.J. Smith, Jr. 1979. Biological control with mycoherbicides. Ann. Rev. Phytopathology 17:301310.
Walker, H.L., and W.L. Connick, Jr. 1983. Sodium alginate for production and formulation of mycoherbicides. Weed Sci. 31:333338.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}